chapter ten
Microminds and
Macrominds
WE RETURN TO THE IMPERFECT but comfortable realm of experience. In chapters
5 and 6, we sought out predictions in hologramic theory. Now we shift emphasis to explanations, to the
use of hologramic theory for making rational sense of certain equivocal and seemingly unbelievable
observations. And we start with the behavior of bacteria.
Bacilli, rod-shaped bacteria, propel themselves through fluid with whip-like
appendages called flagella. flagellar motion depends on a contractile protein
hooked to the base of each microbial appendage. An individual flagellum rotates, in the process executing wave motion. Locomotion of a bacillus, therefore, is an algebraic function of the phase and
amplitude spectra collectively generated by its flagella. We might even regard the overt behavior
of a bacillus as a direct consequence of periodic activity--the phase and amplitude
spectra--reflecting the rhythm of its contractile proteins. Thus if a hologramic mind
exists in the bacillus, it shows up literally and figuratively right at the
surface of the cell.
But is there really a mind in a creature so primitive? We can describe a
stretching and recoiling spring with tensor transformation. But our intuitions would balk,
and rightfully, if we tried to endow the back porch door with hologramic mind. Thus
before we apply hologramic theory to the bacillus, we need evidence only
experiments and observations can supply.
Single cells of many sorts can be attracted or repelled by various chemicals.
The reaction is called chemotaxis. In the 1880s, when bacteria was still a very
young science, a German botanist named Wilhelm Pfeffer made an incredible
observation in the common intestinal bacillus, Escherichia coli (E.
coli)-- more recently of food-poisoning fame. Other scientists had previously
found that meat extract entices E. coli whereas alcohol repels them: the
bacteria would swim up into a capillary tube containing chicken soup or hamburger juice but would avoid one with ethanol. What would happen, Pfeffer wondered if
he present his E. coli with a mixture of attractants and repellants? He found if concentrated enough, meat
juice would attract E. coli even though the concentration of ethanol was
enough, by itself, to have driven the bacteria away.
Did Pfeffer's observations mean that bacteria make decisions? Naturally, the
critics laughed. But in comparatively recent times biochemists have begun to
rethink Pfeffer's question. And in rigorously quantified experiments with
chemically pure stimulants, two University of Wisconsin investigators, J. Adler
and W-W Tso came the conclusion, "apparently, bacteria have a 'data processing'
system." Courageous words, even within the quotations marks. For in a living
organism, 'data processing' translates into what we ordinarily call thinking.
Adler and Tso established to important points. First, the relative--not
absolute--concentrations of attractant versus repellent determines whether E.
coli will move toward or away from a mixture. Second, the organisms do not
respond to the mere presence of stimulants but instead follow, or flee, a
concentration gradient. And it was the consideration of concentration gradient that led
biochemist D. E. Koshland to establish memory in bacteria.
Koshland became intrigued by the fact that a creature only 2 micrometers long
(.000039 inches) could follow a concentration gradient at all. The cell would
have to analyze changes on the order of about one part in ten thousand--the
rough equivalent of distinguishing a teaspoon of Beaujolais in a bathtub of
gin, a "formidable analytical problem," Koshland wrote.
Did the cell analyze concentration variations along its length? To Koshland's
quantitative instincts, 2 micrometers seemed far too short a length for that.
Suppose, instead, the bacterium analyzes over time, instead of space (length)?
What if the cell could remember the past concentration long enough to
compare it with the present concentration? Koshland and company knew just the
experiment for testing between the two alternative explanations.
When a bacillus is not responding to a chemical stimulus, it tumbles randomly
through the medium. (Its flagella crank randomly.) In the presence of a stimulus, though, the bacterium checks
the tumbling action and swims in an orderly fashion. What would happen, Koshland
wondered, if he tricked them? What if he placed the organism in chemical stimulus
(no gradient though) and then quickly diluted the medium? If the bacteria
indeed analyzes head-to-tail, they should go right on tumbling because there'd
be no gradient, just a diluted broth. But if bacteria remember a past
concentration, diluting the medium should fool them into thinking they were in
a gradient. Then they'd stop tumbling.
"The latter was precisely what occurred," Koshland wrote.[1] The bacterial relied on memory
of the past concentration to program their behavior in the present.
There was more. Koshland called attention to another feature of bacterial
behavior. He pointed out that in responding to a chemical stimulus--in
checking the tumbling action-- "the bacterium has thus reduced a complex problem
in three-dimensional migration to a very simple on-off device."[2]
When a human being simplifies a complicated task, we speak about intelligence. Thus bacteria show evidence of rudimentary minds.
And we can use hologramic theory to account for it.
***
Adler and Tso discovered that attractants induce counterclockwise rotation in
E. coli's flagella. Repellents cranked the appendage clockwise. In terms of
hologramic theory in its simplest form, the two opposite reactions are 180
degrees (or pi) out of phase. By shifting from random locomotion to movement
toward or away from a stimulus, the organism would be shifting from random
phase variations in its flagella to harmonic motion--from cacophony to a melody
if they were tooting horns instead of churning their appendages.
Adler and Tso identified the bacterium's sensory apparatus. Like the
biochemical motor, it also turned out to be a protein. A search eventually turned
up strains of E. coli that genetically lack the protein for a specific attractant. (Specific
genes direct the synthesis of specific proteins.)
At the time they published their article, Adler and Tso had not isolated the
memory protein (if a unique one truly exists). But the absence of that information doesn't prevent our using
hologramic theory to explain the observations: Phase spectra must be
transformed from the coordinates of the sensory proteins through those of
contractile proteins to the flagella and into the wave motion of the
propelling cell. Amplitudes can be handled as local constants. The chemical
stimulus in principle acts on the bacterium's perceptual mechanism analogous to
the reconstruction beam's decoding of an optical hologram. As tensors in a
continuum, the phase values encoded in the sensory protein must be transformed
to the coordinate system representing locomotion. The same message passes from
sensory to motor mechanisms, and through whatever associates the two. Recall
that tensors define the coordinates, not the other way around. Thus, in terms
of information, the locomotion of the organism is a transformation of the
reaction between the sensory protein and the chemical stimulus, plus or minus
the effects of local constants. Absolute amplitudes and noise, products of
local constants, would come from such things as the viscosity of the fluid (e.g., thick pus versus sweat) the
age and health of organisms, the nutritional quality of the medium (better to
grow on the unwashed hands of a fast-food hamburger flipper than on the
just-scrubbed fingernails of a surgical nurse), or whatever else the phase spectrum
cannot encode. As for the storage of whole codes in small physical spaces,
remember that phase has no prescribed size in the absolute sense. A single
molecule can contain a whole message.
***
Evidence of memory on single-celled animals dates back at least to 1911, to
experiments of the protozoologists L. M. Day and M. Bentley on paramecia.[3] Day and Bentley put a
paramecium into a snug capillary tube--one whose diameter less than the
animal's length. The paramecium swam down to the opposite end of the tube,
where it attempted to turn abound. But in the cramped lumen, the little fellow
twisted, curled, ducked, bobbed....but somehow managed by accident to get faced
in the opposite direction. What did it do? It immediately swam to the other
end and got itself stuck again. And again it twisted, curled, ducked...and only
managing to get turned around by pure luck. Then, after a while Day and
Bentley began to notice something. The animal was taking less and less time to
complete the course. It was becoming more and more efficient at the tricky
turn-around maneuver. Eventually, it learned to execute the move on the first
attempt.
Day and Bentley's observations didn't fit the conventional wisdom of their day, nor the criteria for learning
among some schools of thought in our own times. Their little paramecia had taught themselves the trick, which in
some circles doesn't count as learning. But in the 1950s an animal
behaviorist named Beatrice Gelber conditioned paramecia by the same basic
approach Pavlov had taken when he used a whiff of meat to make a dog drool
when it heard the ringing of a bell.
Gelber prepared a pâté of her animal's favorite bacteria (a single
paramecium may devour as many as 5 million bacilli in a single day[4] ); then she smeared some of it
on the end of a sterile platinum wire. She dipped the wire into a
paramecium culture. Immediately her animals swarmed around the wire, which was
not exactly startling news. In a few seconds, she withdrew the wire, counted off a few more
seconds and dipped it in again. Same results!. But on the second trial,
Gelber presented the animals with a bare, sterilized wire, instead of with bacteria.
No response! Not at first, anyway. But after thirty trials--two offers of
bacteria, one of sterile wire--Gelber's paramecia were swarming around the
platinum tip, whether it proffered bacterial pâté or not.[5]
Naturally, Gelber had her critics, those who dismiss the idea that a single
cell can behave at all, let alone remember anything. I must admit, a mind isn't easy to
fathom in life on such a reduced scale. Yet I've sat entranced at my stereoscopic microscope for hours on
end watching protozoa of all sorts swim around in the water with my
salamanders. I've often wondered if Gelber's critics had ever set aside their
dogmas and doctrines long enough to observe for themselves the truly remarkable
capabilities of one-celled animals. Let me recount something I witnessed one
Saturday afternoon many years ago.
On occasion, a fungal growth infects a salamander larva's gills. To save the
salamander, I remove the growth mechanically. On the Saturday in question, I
discovered one such fungal jungle teeming with an assortment of protozoa. What
were those beasts? I wondered. Instead of depositing the teased-off mass on
the sleeve of my lab coat, I transferred it to a glass slide for inspection
under the much greater magnification of the compound phase microscope.[6]
Several different species of protozoa were working the vine-like hyphae
of the fungus. I was soon captivated by the behavior of a species I couldn't
identify. They were moving up and down the hyphae at a brisk pace. At the
distal end of a strand an animal's momentum would carry it out into the
surrounding fluid. It would then turn and swim back to its "own" hypha, even
when another one was closer. Something spatial or chemical, or both, must be
attracting these critters, I thought almost out loud. Just as I was thinking
the thought, one animal attracted my attention. It had just taken a wide
elliptical course into the fluid; but along the return arc of the excursion,
another hypha lay directly on its path. And my little hero landed on it.
After a few pokes at the foreign strand, the animal paused as though something
didn't seem quite right. Meanwhile its sibs were busily
working the territory. After a few tentative pokes, my animal moved away. But
now it landed on a third hypha, shoved off after a brief inspection and landed
on still another hypha. Soon it was hopelessly lost on the far side of the
microscopic jungle.
But then something happened. As I was anticipating its departure, protozoan
hesitated, gave the current hypha a few sniffs and began slowly working up and
down the shaft. After maybe five or six trips back and forth along the strand,
my animal increased its speed. Within a few minutes, it was working the new
hyphas as it had been when it first attracted my attention. I couldn't escape
the thought that the little creature had forgotten its old home and had learned
the cues peculiar its new one.
Had I conducted carefully controlled experiments, I might have discovered a
purely instinctive basis for all I saw that Saturday. Maybe Gelber's or Day
and Bentley's observation can be explained by something other than learning,
per se. But, instinctive or learned, the behavior of protozoa--or
bacteria--doesn't fit into the same class of phenomena as the action-reaction
of a rubber band. Organized information exists in the interval between what
they sense and how they respond. We employ identical criteria in linking
behavior to a human mind.
***
But higher organisms require a "real" brain in order to learn, don't they? If
posing such a question seems ridiculous, consider an observation of a
physiologist named G. A. Horridge made some years ago on decapitated roaches
and locusts.
In some invertebrates, including insects, collections of
neurons--ganglia--provide the body with direct innervation, as do the spinal
cord and brainstem among vertebrates. Horridge wondered if ganglion cells
could learn without the benefit of the insect's brain. To test the question,
he devised an experiment that has since become famous enough to bear his name: "The
Horridge preparation."
In the Horridge preparation, the body of a beheaded insect is skewered into an
electrical circuit. A wire is attached to a foot. Then the preparation is
suspended directly above a salt solution. If the leg relaxes and gravity pulls
down the foot, the wire dips into the salt solution, closing the electrical
circuit and-- zappo! A jolt is delivered to the ganglion cells
within the headless body. In time, the ganglion cells learn to avoid the shock
by raising the leg high enough to keep the wire out of the salt solution.
An electrophysiologist name Graham Hoyle went on to perfect and refine the
Horridge preparation. Working with pithed crabs, he used computers to control
the stimuli; he made direct electrophysiological recordings from specific
ganglion cells; and, because he could accurately control the stimuli and record the responses, Hoyle
was able to teach the cells to alter their frequency of firing, which is a very
sophisticated trick. How well did the pithed crabs
learn? According to Hoyle, "debrained
animals learned better than intact ones."[7]
I'm not suggesting that we replace the university with the guillotine. Indeed,
later in this chapter we'll bring the brain back into our story. But first-rate evidence of mind exists in some very hard-to-swallow places. Brain (in the
sense of what we house inside our crania) is not a sine qua non of mind.
Aha, but does the behavior of beheaded bugs really have any counterpart in like us?
***
In London, in 1881, the leading holist of the day, F. L. Goltz arrived from
Strasbourg for a public showdown at the International Medical Congress with his
arch-rival, Englishman David Ferrier who'd gained renown for investgating functional localization within
the cerebral cortex.
At the Congress, Ferrier presented monkeys with particular paralyses following specific ablations of what came to be known as the motor cortex. Ferrier's experiments were so dramatically elegant that he won the
confrontation. But not before Goltz had presented his "Hund ohne Hirn," (dog without brain)-- an animal who
could stand up even though his cerebrum had been amputated. [8]
The decerebrated mammal has been a standard laboratory exercise in physiology courses
every since. A few years ago, a team of investigators, seeking to find out if
the mammalian midbrain could be taught to avoid irritation of the cornea, used the blink reflex to demonstrate
that, "decerebrate cats could learn the conditioned response."[9]
Hologramic theory does not predict that microbes, beheaded bugs or decerebrated
dogs and cats necessarily perceive, remember and behave. Experiments furnish
the underlying evidence. Some of that evidence, particularly with bacteria,
has been far more rigorously gathered than any we might cite for support of
memory in rats or monkeys or human beings. But the relative nature of phase
code explains how an organism only 2 micrometers long--or a thousand times
smaller that, if need be--can house complete sets of instructions.
Transformations within the continuum give us a theory of how biochemical and
physiological mechanisms quite different from those in intact brains and bodies
of vertebrates may nevertheless carry out the same overall informational
activities.
Yet hologramic theory does not force us to abandon everything else we know.[10] . Instead, hologramic theory
gives new meaning to old evidence; it allows us to reassemble the original
facts, return to where our quest began, and with T. S. Eliot, "know the place
for the first time."
In the last chapter, I pointed out that two universes developed according to
Riemann's plan would obey a single unifying principle, curvature, and yet could
differ totally if the two varied with respect to dimension. Thus the
hologramic continuum of both a salamander and a human being depend on the phase
code and tensor transformations therein. But our worlds are far different from
theirs by virtue of dimension. Now let's take this statement out of the
abstract.
***
It's no great surprise to anyone that a monkey quickly learns to sit patiently in front of a
display panel and win peanut after peanut by choosing, say, a triangle instead of a
square. By doing essentially the same thing, rats and pigeons follow the same
trend Edward Thorndike first called attention to in the 1890s. Even a goldfish
when presented with an apparatus much like a gum machine soon learns that
bumping its snout against a green button will earn it a juicy
tubifex worm while a red button brings forth nothing. Do choice-learning
experiments mean that the evolution of intelligence is like arithmetic: add
enough bacteria and we'd end up with a fish or an ape? In the 1950s a man
named Bitterman began to wonder if it was all really that simple. Something
didn't smell right to him. Bitterman decide to add a
new wrinkle to the choice experiments.
Bitterman began by training various species to perform choice tasks. His animals had to
discriminate, say, A from B. He trained goldfish, turtles, pigeons,
rats and monkeys to associate A with a reward and B with receiving nothing.
Then Bitterman played a dirty trick. He switched the reward button. Chaos
broke out in the laboratory. Even the monkey became confused. But as time
went by, the monkey began to realize that now B got you the peanut, not A.
Then the rat began to get the idea. And the pigeon too! Meanwhile over
in the aquarium, the goldfish was still hopelessly hammering away at the old
choice. Unlike the other members of the menagerie, the fish could not kick its
old habit and learn a new one.
What about his turtle? It was the most interesting of all Bitterman's
subjects. Confronted with a choice involving spatial discrimination, the
turtle quickly and easily made the reversal. But when the task involved visual
recognition, the turtle was as bad the goldfish; it couldn't make the switch.
It was as though the turtle's behavior lay somewhere between that of the fish
and the bird. Turtles, of course, are reptiles. During vertebrate evolution,
reptiles appeared after fishes (and amphibians) but before birds and mammals.
Now an interesting thing takes place in the evolution of the vertebrate brain. In
reptiles, the cerebrum begins to acquire a cortex on its external surface. Was
the cerebral cortex at the basis of his results? Bitterman decided to find out
by scaping the cortex off the rat's cerebrum. Believe it or not, these rats successfully reversed the habit when give a
space problem. But they failed when the choice involved visual discrimination.
Bitterman's rats acted like turtles!
***
Bitterman's experiments illustrate that with the evolution of the cerebral
cortex something had emerged in the vertebrate character that had not existed
before. Simple arithmetic won't take us from bacterium to human being.
As embryos, each of us re-enacts evolution, conspicuously in appearance but subtly in behavior, as well. At first we're more like a slime mold than an animal. Up to about the fourth interuterine month, we're quite
salamander-like. We develop a primate cerebrum between the forth and sixth
month. When the process fails, we see in a tragic way how essential the human
cerebral cortex is to the "Human Condition," in the Barry N. Schwartz sense of of the term.
Mesencephalia is one of the several terms applied to an infant whose
cerebrum fails to differentiate a cortex.[11] A mesencephalic infant
sometimes lives for a few months. Any of us (with a twist of an umbilical
cord, or if mom took too long a swig on a gin bottle) might have suffered the
same fate. Like its luckier kin, the mesencephalic child will cry when
jabbed with a diaper pin; will suckle a proffered breast; will sit up; and it can
see, hear, yawn, smile and coo. It is a living organism. But human though its
genes and chromosomes and legal rights may be, it never develops a human personality, no matter
how long it lives. It remains in the evolutionary bedrock out of which the
dimensions of the human mind emerge. It stands pat at the stage where the human
embryo looked and acted like the creature who crawls from the pond.
Yet there's no particular moment in development when we can scientifically delineate human
from nonhuman: no specific minute, hour or day through which we can draw a neat
draftsman's line. Development is a continuous process. The embryo doesn't
arrive at the reptilian stage, disassemble itself and construct a mammal, de
novo. In embryonic development, what's new harmoniously integrates with what's
already there to move up from one ontological step to the next.[12] The embryo's
summations demand Riemann's nonlinear rule: curvature!
What is arithmetic, anyway. What do we mean by addition and subtraction? At
the minimum, we need discrete sets. The sets must equal each other--or be
reducible to equal sets be means of constants; and their relationships must be
linear. The correct adjective for describing a consecutive array of linear set
is contiguous (not continuous), meaning that the successive members
touch without merging into and becoming a part of each other--just the opposite
of Riemann's test of continuity. This may seem utterly ridiculous. But if the
sets in 1 + 1 + 1 surrender parts of themselves during addition, their sum
will be something less than 3. We literally perform addition and subtraction
with our fingers, abacus beads, nursery blocks and digital computers-- any
device that uses discrete, discontinuous magnitudes.
Continuity is essential to the theory of evolution. Try to imagine a tree
whose branches are not and never were continuous, back to the main trunk.
Continuity in fact during embryonic development is our prima facie
evidence for continuity in theory among the species. Evolution is
inconceivable in a simple, discontinuous arithmetic system. In the light of
Bitterman's turtle, a straight-line theory of the natural history of
intelligence would predict discontinuity among the species and render the
theory of evolution itself no more defensible on formal grounds than the events
depicted in Genesis. Bitterman's investigations deny a simple, linear
progression from fish to human and provide experimental evidence for the
evolution of intelligence.
We have not constructed hologramic theory along linear lines. If we had, we
wouldn't be able to reconcile what we find. We would be forced to ignore some
facts in order to believe others. Without the continuum, we would be unable to
explain not only the differences and similarities of the species, but also
those in ourselves at various stages of our own embryonic development.
The hologramic continuum, by nature, allows new dimensions to integrate
harmoniously with those already present. It lets us explain how our biological
yesterday remains a part of today within a totally changed informational
universe. Even though we share the same elemental rule--the phase code--with
all other life forms, we're not reducible to what we once were, nor to bacteria
or beheaded bugs. We are neither a linear sum of what we were nor a linear
fraction of what we used to be. And our uniquely human inner world begins to
unfold with the advent of the cerebral cortex.
***
Physiologically, the cerebral cortex was a near-total enigma until
comparatively recent times. But two physiologists, David Hubel and Torsten
Wiesel spent decades exploring pattern recognition in the visual cortex, first
of cats and eventually monkeys.[13] They identified three basic
types of neurons there, cells that would respond to visual targets of different
degrees of complexity. Hubel and Wiesel called these neurons simple,
complex and hypercomplex cells:
- Simple cells fired in response to
a barlike image in a fixed position with a particular orientation; those that
would fire in response to, say, a horizontal target would quite immediately if
the bar were tilted or if Hubel and Wiesel substituted it with a vertical or
bar.
- Complex cells were those that went on firing to a basically
two-dimensional target. But the complex cells would not fire if the target's
pattern became still more complicated.
- That's where hypercomplex cells come
into the picture: they were cells that kept on discharging when complex cells
stopped.
The visual system must handle a great deal of information in addition to
patterns: it must deal with color, motion, distance, direction--with a variety of independent
abstract dimensions that have to be compiled into a single, composite picture, as do bars and rectangles and edges to make unified percept.
Hubel and Wiesel analogized the problem to a jigsaw puzzle: the shape of the
pieces is independent of the picture or potential picture they bear. We recognize a
checkerboard whether it's red and black or black and blue. And we don't usually
confuse a Carmen-performing diva flitting around the stage in a red and
black dress with the red-black checkerboard.
We're always assembling informational dimensions into a single composite scene.
If we go to a three-dimensional object, or if the object is moving, or if we
attach some emotional significance to the input, we must either integrate the
data into a percept or keep the subsets sorted into groups. And the
integration (or segregation) must be quick. Vary the beast's capability for
handling dimensions and we change its perception in a nonlinear way, as
Bitterman did when he took the knife to his rat's cortex. Interestingly, it
was not until Hubel and Wiesel began studying the monkey (versus the cat) that
they discovered hypercomplex cells in sufficient numbers to analyze their
complicated physiological details.
What we hear, touch, taste and smell may also be multidimensional. For
instances, we may recognize a melody from The Barber of Seville, but our
understanding of the lyrics may depend on a knowledge of Italian. And whether
we're enthralled or put to sleep will depend on factors other than just the
words or the music.
We also harmoniously integrate diverse sensory data. Thus silent movies
disappeared quickly after talkies came along. We not only have the capacity to
combine sight and sound, but we like (or hate) doing it which is a whole other
constellations of dimensions.
The cerebral cortex is far from the only dimensional
processor in the brains of organisms (although it may very well be the most
elegant one in there). The frog, for instances, has a well developed roof
on its midbrain--the tectum I mentioned in chapter 3: the part that helps
process visual information among non-mammalian vertebrates. While the tectum
comes nowhere close to the capabilities of a primate visual cortex, it
nevertheless integrates different dimensions of the frog's visual perception.[14]
***
Another structure I mentioned in chapter 3 was the zucchini-shaped
hippocampus, lesions of which induce short-term memory defects and
make it difficult for persons so afflicted to repeat newly presented phrases or
sequences of numbers. In rats, however, the hippocampus appears to assist
navigation.
The function of the rat's hippocampus became evident in studies involving what
is called the radial maze.[15]
This kind of apparatus consists of typically a dozen alleys leading off a
central choice point like the spokes radiating from the hub of a wagon wheel.
To win a reward, the rat must go through the alleys in a predetermined sequence. To carry out the task, the rat must remember
which alley he's in and which turn to take for the next one. The rat must
compile at least two sets of memories: one of positions, the other of
direction. Lesions in the hippocampus erode the animal's efficiency at the
radial maze.
Now think of what happens when we recite the lines of a poem. We must remember
not only the individual words but, so as to place correct emphases, their
location in the sequence.[16]
Although the task assumes verbal form in a human being, its informational
aspects seem quite like those a rat uses to organize memories of geographic
locations and sequences.
At the same time, clinical and laboratory evidence don't prove that the
hippocampus is the exclusive seat of such short-term memory processing.
Lesions do not totally nullify the rat's ability to run the radial maze:
performances dropped from 7 correct turns out of 8 to 5 or 6, a statistically
significant drop but far from the total loss of the ability that follow from
destruction of the seat of the neural information. [17] I once watched a record film
of a man with a damaged hippocampus. He made errors when repeating phrases,
but he wasn't always wrong. In addition, he often employed subtle tricks to
recall items. When he was allowed to count on his fingers, he could often
correctly repeat phrases that he couldn't handle without them. Other parts of the brain can compile memories of position and distance but
would seem to do with much less efficiency than the hippocampus.
The success or failure of a particular behavior may depend on how fast an
organism can assemble different memories. Out in the wild, the navigational
problems a rat confronts are much more difficult--and potentially
perilous--than anything in the laboratory. Ethologist are students of behavior
in the wild. And ethologists Richard Lore and Kevin Flanders, undertook the
frightening job digging up a rat-infested garbage dump in New Jersey to see how
the beasts live out there. To Lore and Flander's surprise, they found that
wild rats live in family groups, each with its own burrow. The dump
wasn't honeycombed with one communal rat flop, the animals randomly infesting a
labyrinth and eating, sleeping or mating wherever the opportunity presented
itself. Now the wild rat is one vicious creature, as a child of the inner city
can often testify first hand. A strange rat who ends up in the wrong hole
isn't welcomed as an honored dinner guest but may very well become the piece de
résistance. Thus when the rat ventures into the night and turns
around to bring home a half-eaten pork chop, it by-god better know which burrow
to choose out of hundreds in a multidimensional array. It would quickly
succumb to its own social psychology if not for the superb navigational system
resident in its hippocampus.
The use to which a rat puts its hippocampus seems at least as complicated as ours.
Appreciate, though, that while the qualitative features of the behaviors
mediated by the homologous brains structures can differ greatly between them and
us, the abstract attributes--the analogous logic-- can be quite similar. Let's
remind ourselves of this from hologramic theory.
We can construct two continua that have identical numbers of dimensions yet
produce different universes. How? The shape of a universe depends not only on
how many dimensions it has, but on how they connect up and which part connects with what. Recall in the last
chapter we envisaged adding a dimension by converting a figure 8 into a
snowman.
What if we take a snowman apart and reassemble it with the members in a
different order. Call our initial snowman A and our reshuffled one B. Now
let's send an imaginary insect crawling on each figure at the same time and at
the same speed. Of course, both insects will complete the round-trip excursion
together. But relative to the starting point (the phase variation between the two), the two bugs will never once
have traveled an equal distance until both have arrived back at the start-finish point. The
similarities and differences don't trick us. Likewise, the differences
between our hippocampus and the rat's shouldn't mask the similarities.
As I mentioned earlier, some psychologists conceptualize short-term or working
memory by means of an idealized compartment.[18] The hippocampus, as I've
suggested, may be a location of a portion of that compartment, at least in
higher animals. Of course, as evidence from, for instance, decerebrated cats
and many other sources indicates, we can't consider the hippocampus as the
exclusive repository of short-term memory. But hippocampal functions may yet
reveal critical details about the dynamics of perception and reminiscence in
higher animals. Circumstantial evidence from rats as well as humans indicates
that an active memory in the hippocampus is short term. Now short-term memory
in general is very sensitive to electroconvulsive shock (ECS). And relatively
mild electrical stimulation of the hippocampus or the surrounding brain can
evoke a violent convulsive seizure.[19]
A temporary and erasable working memory would be valuable to us as well as to rats. When we no longer
need a telephone number, for example, we simply expunge. Yet we wouldn't want
to forget all telephone numbers. After a journey from the garbage pile, the
neural map in the rat's hippocampus could become a liability. But the rat
wouldn't want to have to relearn every map. ("That's the pipe were the coyote
got my brother!") What might control the shift from short-term to long-term
storage? I raise the latter question not to signal a final answer. But a little
speculation from a few facts will allow me to illustrate how we can use
hologramic theory to generate working hypotheses (the kind from which
experimentation grows). Also, the hippocampus exhibits an important feature of the brain that Norbert Wiener predicted many years ago.
The human hippocampus interconnects with vast regions of the central nervous system. Its most conspicuous pathways lead to and from subdivisions of what is called the limbic system. The limbic system is most conspicuously
associated with emotions. 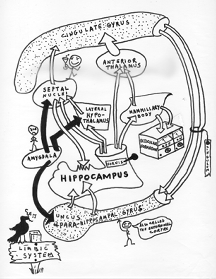 One hippocampal circuit in particular connects the
hippocampus to a massive convolution called the cingulate gyrus. Draped like a lounging leopard on the corpus callosum, the cingulate gyrus was the first part of the limbic system ever designated [20] and became a favorite target of
psychosurgery in the post-prefrontal lobotomy era.
The hippocampal-cingulate circuit has (among other many things) three features
that are highly germane to our discussion.
One hippocampal circuit in particular connects the
hippocampus to a massive convolution called the cingulate gyrus. Draped like a lounging leopard on the corpus callosum, the cingulate gyrus was the first part of the limbic system ever designated [20] and became a favorite target of
psychosurgery in the post-prefrontal lobotomy era.
The hippocampal-cingulate circuit has (among other many things) three features
that are highly germane to our discussion.
- First, a number of relay stations intervene between the hippocampus
and the cingulate gyrus. These relay stations are locations where the signal
can be modified; where messages can be routed to and from other areas of the
brain and spinal cord and blended (or removed) with (from) other data.
- Second, the circuit consists of parallel pathways (co- and
multi-axial) all the way around. The significance of this is that when
activated the circuit preserves phase information in principle in the way
Young and Fresnel did when making interference patterns--by starting from the
same source but varying the specific course between the referencing waves.
The hippocampal-cingulate circuitry looks designed to handle phase
differentials, and on a grand scale!
- Third, the overall circuit makes a giant feedback loop. Feedback is
at the heart of the communications revolution Norbert Wiener touched off in
1947 with the publication of his classic, Cybernetics and that you're living through as you read these very words.
***
Biofeedback became a popular subject a couple of decades ago when people were
hooking electrodes to the their heads and stopping an starting electrical
trains by altering their thoughts. "EEG with electric trains," a friend of
mine used to call what seemed to him a stunt wherein amplified electrical
signals from the scalp are fed into the transformer of an electric train set
instead of a polygraph. However one wants to characterize feedback, it is
central to much that goes on chemically and physiologically in our bodies--including
the brain.
Wiener spent World War Two trying to develop methods for anticipating where a
German bomber was headed so that an Allied aircraft gun crew could aim
properly to shoot it down, evasive actions and the trembling ground notwithstanding. The problem led
Wiener began to appreciate the importance of the feedback loop in controlling the
output from a system where the output device itself is subject to continual
changes. Feedback became the central notion in his cybernetic theory.
What a feedback loop does is relate input to output in a dynamic instead of
static way. With moment-to-moment monitoring, tiny corrections can be made in
the output to compensate for last minute changes not only in a target but also
in the readout mechanisms.
Wiener the mathematician used to joke about his
ignorance of the brain. Nevertheless, in 1947 from pure cybernetic
theory, he predicted one of the brain's (or computer's) most important functional attributes:
reverberating circuits. (Reference to "reverberating circuit" is to be found
in even standard textbooks.[21]
) Even though he'd never even seen the cerebellum before, he predicted both
its function and the effects of damage to it (dysfunction in dynamic motor
control).[22]
In a system like that conceived by Wiener, a short-term memory would
reverberate around the feedback loop and remain active until other input
modified it or shut it off. The hippocampus seems elegantly designed for just
that kind of activity.
The specific types of memories associated with the human hippocampus are
especially susceptible to emotions. Tell a class of students that a
particular fact will appear on the final examination and it's almost as though
their limbic systems suddenly opened the gates to the permanent storage
compartment. On the other hand if you become scared or angry or amorous just
before you dial a newly looked-up telephone number, you'll probably have to
consult the directory again before putting in the call.
Karl Pribram has suggested that neural holograms exist in microcircuits within
neurons. A cogent argument can be made for his thesis, but not in the
short-term memory on the hippocampus. The evidence suggests that
short-term hippocampal memory depends on a vast number of neurons, perhaps even the
entire feedback loop. Let me repeat an important maxim of hologramic
theory: the theory won't predict the biochemical or physiological mechanisms of
memory; the absolute size of a whole phase code is arbitrary, meaning that it
may be (but doesn't have to be) tiny, as Pribram's microcircuits, or in
oscillations within a pair of molecules on a cell membrane; or gigantic as in
the entire hippocampus, or even with the whole nervous system. Recall that the
same code may exist simultaneously in many different places or sizes and in
many specific mechanisms. To make the memory of a telephone number permanent,
what has to transform is not a protein or a voltage but a set of variational
relationship--the tensors of our hologramic continuum.
In 1922, Einstein wrote The Meaning of Relavity. In it, he demonstrates
that his special relativity theory (E=mc2) can be derived from the
tensors of his general relativity theory, from his four-dimensional space-time
continuum. Although the hologramic continuum is not the same as Einstein's
theory, we do use Riemann's ideas, as did Einstein. The logic is similar.
Einstein demonstrated the same relative principle in both the very small atom
and in the universe at large. Hologramic theory, too, tells us that the same
elemental rules operate in mind in the small or the large. If we find a
memory in, say, the resonance of two chemical bonds within a hormone molecule,
we should not throw up our hands in disgust abandon science for magic simply because someone else discovers
the same code in the entire feedback loop of the hippocampus. Thus if
microcircuits turn out to play no role in the short-term memory on the
hippocampus, they could still very
well exist elsewhere.
***
The cortex on the cerebrum's frontal lobe may also be essential to the uniquely
human personality, and some of the data collected with reference to it are not
only useful to our discussion but also rather interesting in their own right.
One of the most important contributors to knowledge about the frontal lobe is no less than Karl
Pribram.
Human beings who have undergone prefrontal lobotomy often cannot solve problems
consisting of many parts. Pribram, who was familiar with these clinical signs
as a neurosurgeon, had a hunch about why, a hunch he pursued in the lab as a
neurophysiologist.
Signals from the frontal lobes often have an inhibitory effect on other areas
of the brain. (One theory behind prefrontal lobotomy was that it relieved the
uptights.) Pribram began wondering if such inhibitory activity might represent
something akin to parsing a message-- breaking a sequence of letters or words
into meaningful chunks. Maybe to lobotomized patients words such as
"hologramic theory" seem like "ho logramichteory"; or "ho log ram ich teory."
He tested his hypothesis on monkeys with what he called "the alternating
test," a modified version of the shell game as he describes it.[23]
In the alternating test, the monkey sits in front of two inverted cups, one
with a peanut under it, the other not. To win the peanut, the monkey must turn
up the cup opposite the winning choice on the last trial (thus the name
'alternating'). At the end of a trial a screen descends, blocking the
monkey's view of the cups and remaining down anywhere from a few seconds to
several hours. Monkeys find the game easy, and, after a little experience,
readily win peanuts, even after the screen has been down all day long. But
following frontal lobotomy, Pribram found the monkey "will fail at this simple
task even when the interval between trials is reduced to three seconds."[24]
It occurred to Pribram that "perhaps the task appears to these monkeys much as
an unparsed passage does to us." What would happen, he wondered, if he
organized the message for the lobotomized monkey? He thought he could do
the parsing by alternating short and long pauses between trials. In the
original test the interval between trials was randomly selected (for
statistical prudence). Now, though, the animal would sit in front of the cups,
as before, and would again win a peanut by selecting the cup opposite the
correct choice on the previous trial. But the screen would remain down for
either 5 or 15 seconds. Alternating short and long pauses was to akin to
converting an amorphous sequence, as for example LRLRLRLR, into sets like (L+R)
(L+R) (L+R), the 5-second pauses representing the plus signs and the 15-second
pauses the parentheses.
How did the lobotomized monkeys make out? In Pribram's words, "immediately the
monkeys with frontal cortex damage performed as successfully as the control
animals whose brains were intact."[25]
Pribram's findings have practical implications that go beyond far beyond the
frontal lobe. It might be possible to develop strategies to assist a damaged
brain in carrying out functions it has only apparently lost. Such clinical
strategies could conceivably evolve from exercises like finding the common
denominator in the functions of the rat versus human hippocampi.
***
Primates seem to parse information rather well in comparison to other highly
intelligent organisms. The forward growth of the cerebrum, a hominoid
characteristic, would seem to coincide with parsing capabilities. But
potential pitfalls await us if we oversimplify any brain function. Consider this
anecdote Alexander Luria relates about the English neurologist Sir William
Gower.
Gower had a patient with a speech aphasia that surfaced when the person tried
to repeat certain words, 'no' being one of them. During one session, after
several failed attempts to repeat Gower's sentence, the patient became
exasperated and declared: "No doctor, I can't say 'no.'"[26]
It was as though Gower's patient housed at least two disconnected mental
universes. (The observation seems akin to split brain.[27]) Obviously, if the two
independent universes can't communicate, they can't integrate (or parse, for
that matter). The psychologist, Julian Jaynes[28] has proposed that human
consciousness evolved, and history dawned en passant, following the development
of new connections between the left and right temporal lobes[29]. A capacity for acute
self-awareness, Jaynes cogently argues, is a cardinal characteristic of
present-day human beings. According to Jaynes, when a prehistoric person
reported, for instance, hearing the voice of the gods, he or she probably heard
himself or herself without recognizing the actual source. Our ancestors,
in the Jaynes's theory, had "bicameral" minds--minds like the U. S.
Congress divided into a Senate and House.
I don't know if Jaynes is right or wrong. (And I have no advice as to how to
test his hypothesis experimentally.) But the harmonious blending of
mentalities, like Buster and animals in the looking up-experiments, is
consistent with the hologramic principle of independence--but on a cosmic scale. The
smooth, continuous blending of one universe with another is something the
hologramic continuum can do very easily, in theory. Nor would we need a lobotomy to separate dimensions; that could be done with signals producing the equivlance of desgructive interference--jamming!
***
In chapter 8, when we were optically simulating calculations, I demonstrated
that identical phase-dependent memories can be produced both a priori and a
posteriori. Thus one principle we can infer directly from hologramic theory
is that the ancient argument between rationalists and empiricists (i.e.,
whether ideas are innate or learned) is a phony issue in the quest of
mind-brain. On the one hand, we can use learning rates to evaluate memory from
but can't define or reduce the hologramic mind to learning and experience.
Conversely, the question of whether nature or nurture or a combination of both
create(s) a given memory is something we have to establish from empirical
evidence, not a priori with our theory. Yet the empirical evidence
contains many surprises that often appear contradictory and preposterous in
the absence of a theoretical explanation. Let's consider two examples:
language and social behavior.
***
Until about thirty years ago, nothing seemed less innate and more dependent on
experience than the grammars of human languages. Yet one
fundamental tenet of contemporary theoretical linguistics is that all human
languages develop from common, universal, a priori rules of syntax.[30] Since many specific
languages and cultures have emerged and developed in total isolation from
others, yet exhibit common rules, those rules must be present at birth, so the
reasoning goes.
If we accept the latter idea, does it necessarily follow that the genes determine grammar? Maybe. But in terms of constructing a phase code, there's
at least one other possibility: imprinting during interuterine life. It this
sounds wacky, consider the fact that the behavior of ducklings can be
profoundly influenced by sounds they hear while still unhatched, in the egg.[31]
We, as fetuses, receive sonar vibrations set up by our mother's voice and her
heartbeat. The rhythm of the heart and the resonance of the human vocal
apparatus share common features independent of culture, and the modulating
effects of the human body would be the same whether Mom lives on Madagascar or
Manhattan. Now, I'm not asserting that this heart-beat hypothesis is correct.
Hologramic theory really doesn't provide the answer. But it would be very
useful to know if interuterine experiences can set the stage for the
development of language. Science aside, imagine what a poet could do with the
idea that language has its genesis not in our embryonic head but in the hearts
of our original host.
***
A sizable and controversial body of literature exists on the language
capabilities of gorillas and chimpanzees. [32] Some critics of the idea
define human language in such a way that, by definition, the sign language and
symbolic capabilities exhibited by apes are 'behavior,' not 'language.'
Hologramic theory won't resolve the controversy. But if a gorilla and a man
encode and transform the same phase spectrum, they hold a similar thought,
whether its the sensation of an itchy back or the statement, "I'm famished!"
Still, if we invoke the hologramic principle of dimension and also admit the existence of local
constants, we would not reduce the ape's and the man's behaviors to "the same
thing."
***
Turning to social behavior, nothing seemed less learned and more instinctive to the experts of a
generation ago than the social behaviors of nonhuman animals. Herds,
coveys, packs, bands, prides, schools....tend to exhibit rigid, stereotypic
order, showing the same attributes today as when the particular strain of the
species first evolved. But much evidence of social ambiance within animal groups
has come to the fore, especially since Jane Goodall's work with chimps and George Schaller's studies of mountain gorillas put ethology on the map in the 1960s.[33] One study I'm particularly fond, by an ethologist named Gray Eaton, involved a troop of Japanese monkeys.
Eaton
had relocated from southern Japan to a large fenced-in field in Oregon.
Social behavior in these animals is highly structured with a dominance order
among females as well as males. At the top of the entire troop is the
co-called alpha male, which a collaborator of mine, Carl Schneider, use to
call, "the Boss."
What makes for the Boss? Is he the monkey with the sharpest
fangs, quickest fists, meanest temperament, highest concentration of blood
testosterone?
In Eaton's troop, the Boss was a monkey known as Arrowhead. And nobody messed
Arrowhead. Nobody!
Yet Arrowhead was a one-eyed, puny little guy who would have had a tough time
in a fight with most females, let alone any of the other alphas. Eaton even
found lower blood levels of male sex hormone in Arrowhead than among some of
his subordinates. Nor was he aggressive; didn't strut around displaying his
penis; didn't beat up on other monkeys to show everybody else who was in
charge. The troop didn't respect Arrowhead's primacy because of a machismo he
really didn't have but because of a highly complex interplay of social
behaviors and group learning! And the circumstantial evidence suggested that his position in the troop was an outcome of the females' dominance order. The play of baby monkeys often turns into furious if
diminutive combat. Now mother monkey is highly protective. Howls for help
from two small disputants usually bring two angry females head-to-head. Of
course, the weaker mother usually takes her little brat by the arm and scrams,
leaving the field to the stronger female and her infant. Monkeys with weak
mothers grow up learning to run away. Of course, the male monkey with the
dominant mother grows up expecting others to yield to his wishes, which they
tend to do.
Maternalism doesn't end as the male monkey matures. Eaton described Red Witch,
who help establish her grown son as the second-ranking member of the troop.
The son had challenged the second-ranking male who was the tougher the two.
But when her son cried out for help, Red Witch came running. She jumped into the fight and together they
established sonny as number two monkey.
Curiously, Red Witch didn't make her son the Boss. Scrawny Arrowhead was no
match for her. The study didn't really answer the question. But the prospects challenging
Arrowhead seemed like one of those critical taboos that, violated in a
human society, often leads to downfall of the community.
Look at it like this. The job of the Boss is to lead the troop from imminent danger. He must know
the direction from which a hungry leopard may suddenly appear and the quickest
possible escape route. Should a threat come, the Boss must direct an orderly
and efficient retreat: mothers and infants first, he and his hefty lieutenants
last, to put up a fight and, if necessary, die to save the troop (and their
gene pool). Somewhere in the intricate behavior of the group, even Red Witch
leaned not to mess with the Boss of her troop.
***
Jane Goodfield is an historian who has raised insightful questions about
science and scientific ideas. "Why, with very few exceptions," she once wrote,
"have these themes or these people never stimulated great works of literature
or art?"[34] She went on to
observe, "somehow sciences manages to extract the warmth and beauty from the
world."[35]
June Goodfield's words make me feel guilty, and I wish I could deny her
contention. But I can't. And I used to wonder if hologramic theory would so
perfect our understanding of mind-brain as to let no place in the human psyche
for art. Has science finally claimed the last major mystery in Nature? Is
mind now fated to become perfected and boring -- and dehumanized?
I'm not really sure. But I began to phrase the question in terms of what
hologramic theory suggests about intelligence. The result was the final two
chapters of this book.
RETURN TO CONTENTS PAGE
Internet contact:pietsch@indiana.edu